
The cell membrane consists of phospholipids, cholesterol, and proteins, with oligosaccharide chains covalently linked to many of the phospholipid and protein molecules. This limiting membrane functions as a selective barrier regulating the passage of materials into and out of the cell and facilitating the transport of specific molecules. One important role of the cell membrane is to keep constant the ion content of cytoplasm, which differs from that of the extracellular fluid. Membranes also carry out a number of specific recognition and signaling functions, playing a key role in the interactions of the cell with its environment.
Although the plasma membrane defines the outer limit of the cell, a continuum exists between the interior of the cell and extracellular macromolecules. Certain plasma membrane proteins, the integrins, are linked to both the cytoskeleton and ECM components and allow continuous exchange of influences, in both directions, between the cytoplasm and material in the ECM.
Membranes range from 7.5 to 10 nm in thickness and consequently are visible only in the electron microscope. The line between adjacent cells sometimes seen faintly with the light microscope consists of plasma membrane proteins plus extracellular material, which together can reach a dimension visible by light microscopy.
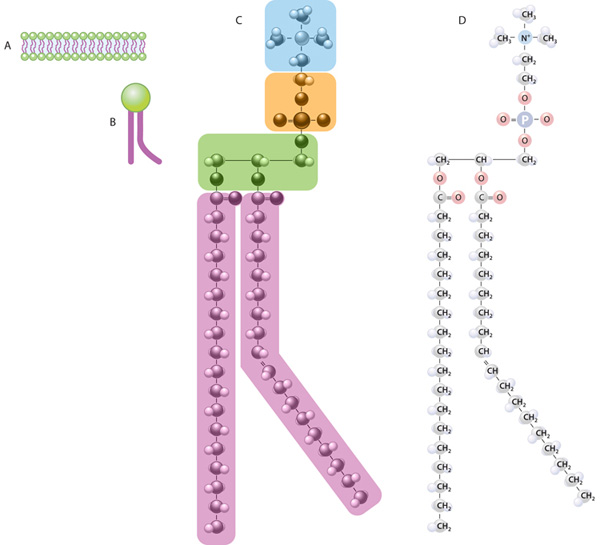
Membrane phospholipids are amphipathic, consisting of two nonpolar (hydrophobic or water-repelling) long-chain fatty acids linked to a charged polar (hydrophilic or water-attracting) head that bears a phosphate group. Phospholipids are most stable when organized into a double layer (bilayer) with the hydrophobic fatty acid chains located in a middle region away from water and the hydrophilic polar head groups contacting the water (Figure 2–1b). Molecules of cholesterol, a sterol lipid, insert at varying densities among the closely-packed phospholipid fatty acids, restricting their movements and modulating the fluidity of all membrane components. The phospholipids in each half of the bilayer are different. For example, in the well-studied membranes of red blood cells phosphatidylcholine and sphingomyelin are more abundant in the outer half, while phosphatidylserine and phosphatidylethanolamine are more concentrated in the inner layer. Some of the outer layer’s lipids, known as glycolipids, include oligosaccharide chains that extend outward from the cell surface and contribute to a delicate cell surface coating called the glycocalyx (Figures 2–1b and 2–2). With the transmission electron microscope (TEM) the cell membrane—as well as all cytoplasmic membranes—may exhibit a trilaminar appearance after fixation in osmium tetroxide; osmium binds the polar heads of the phospholipids and the oligosaccharide chains, producing the two dark outer lines that enclose the light band of osmium-free fatty acids (Figure 2–1b).

(a) Membranes of animal cells have as their major lipid components phospholipids and cholesterol. A phospholipid is amphipathic, with a phosphate group charge on the polar head and two long, nonpolar fatty acid chains, which can be straight (saturated) or kinked (at an unsaturated bond). Membrane cholesterol is present in about the same amount as phospholipid.
(b) The amphipathic nature of phospholipids produces the bilayer structure of membranes as the charged (hydrophilic) polar heads spontaneously form each membrane surface, in direct contact with water, and the hydrophobic nonpolar fatty acid chains are buried in the membrane’s middle, away from water. Cholesterol molecules are also amphipathic and are interspersed less evenly throughout the lipid bilayer; cholesterol affects the packing of the fatty acid chains, with a major effect on membrane fluidity. The outer layer of the cell membrane also contains glycolipids with extended carbohydrate chains.
Sectioned, osmium-fixed cell membrane may have a faint trilaminar appearance with the transmission electron microscope (TEM), showing two dark (electron-dense) lines enclosing a clear (electron-lucent) band. Reduced osmium is deposited on the hydrophilic phosphate groups present on each side of the internal region of fatty acid chains where osmium is not deposited. The “fuzzy” material on the outer surface of the membrane represents the glycocalyx of oligosaccharides of glycolipids and glycoproteins. (X100,000)

(a) The fluid mosaic model of membrane structure emphasizes that the phospholipid bilayer of a membrane also contains proteins inserted in it or associated with its surface (peripheral proteins) and that many of these proteins move within the fluid lipid phase. Integral proteins are firmly embedded in the lipid layers; those that completely span the bilayer are called transmembrane proteins. Hydrophobic amino acids of these proteins interact with the hydrophobic fatty acid portions of the membrane lipids. Both the proteins and lipids may have externally exposed oligosaccharide chains.
(b) When cells are frozen and fractured (cryofracture), the lipid bilayer of membranes is often cleaved along the hydrophobic center. Splitting occurs along the line of weakness formed by the fatty acid tails of phospholipids. Electron microscopy of such cryofracture preparation replicas provides a useful method for studying membrane structures. Most of the protruding membrane particles seen (1) are proteins or aggregates of proteins that remain attached to the half of the membrane adjacent to the cytoplasm (P or protoplasmic face). Fewer particles are found attached to the outer half of the membrane (E or extracellular face). Each protein bulging on one surface has a corresponding depression (2) on the opposite surface.
Proteins are major constituents of membranes (~50% by weight in the plasma membrane). Integral proteins are incorporated directly within the lipid bilayer, whereas peripheral proteins are bound to one of the two membrane surfaces, particularly on the cytoplasmic side (Figure 2–2). Peripheral proteins can be extracted from cell membranes with salt solutions, whereas integral proteins can be extracted only by using detergents to disrupt the lipids. The polypeptide chains of many integral proteins span the membrane, from one side to the other, several times and are accordingly called multipass proteins. Integration of the proteins within the lipid bilayer is mainly the result of hydrophobic interactions between the lipids and nonpolar amino acids of the proteins.
Freeze-fracture electron microscope studies of membranes show that parts of many integral proteins protrude from both the outer or inner membrane surface (Figure 2–2b). Like those of glycolipids, the carbohydrate moieties of glycoproteins project from the external surface of the plasma membrane and contribute to the glycocalyx (Figure 2–3). They are important components of proteins acting as receptors, which participate in important interactions such as cell adhesion, cell recognition, and the response to protein hormones. As with lipids, the distribution of membrane polypeptides is different in the two surfaces of the cell membranes. Therefore, all membranes in the cell are asymmetric.

Both protein and lipid components often have covalently attached oligosaccharide chains exposed at the external membrane surface. These contribute to the cell’s glycocalyx, which provides important antigenic and functional properties to the cell surface. Membrane proteins serve as receptors for various signals coming from outside cells, as parts of intercellular connections, and as selective gateways for molecules entering the cell.
Transmembrane proteins often have multiple hydrophobic regions buried within the lipid bilayer to produce a channel or other active site for specific transfer of substances through the membrane.
Studies with labeled membrane proteins of cultured cells reveal that many such proteins are not bound rigidly in place and are able to move laterally (Figure 2–4). Such observations as well as data from biochemical, electron microscopic, and other studies showed that membrane proteins comprise a moveable mosaic within the fluid lipid bilayer, the well-established fluid mosaic model for membrane structure (Figure 2–2a). Unlike the lipids, however, lateral diffusion of many membrane proteins is often restricted by their cytoskeletal attachments. Moreover, in most epithelial cells tight junctions between the cells (see Chapter 4) also restrict lateral diffusion of unattached transmembrane proteins and outer layer lipids, producing different domains within the cell membranes.
Experiment demonstrating the fluidity of membrane proteins.

(a) Two types of cells were grown in tissue cultures, one with fluorescently labeled transmembrane proteins in the plasmalemma (right) and one without.
(b) Cells of each type were fused together experimentally into hybrid cells.
(c) Minutes after the fusion of the cell membranes, the fluorescent proteins of the labeled cell spread to the entire surface of the hybrid cells. Such experiments provide important data supporting the fluid mosaic model. However, many membrane proteins show more restricted lateral movements, being anchored in place by links to the cytoskeleton.
Membrane proteins that are components of large enzyme complexes are also usually less mobile, especially those involved in the transduction of signals from outside the cell. Such protein complexes are located in specialized membrane patches termed lipid rafts with higher concentrations of cholesterol and saturated fatty acids which reduce lipid fluidity. This together with the presence of scaffold proteins that maintain spatial relationships between enzymes and signaling proteins allows the proteins assembled within lipid rafts to remain in close proximity and interact more efficiently.
The plasma membrane is the site where materials are exchanged between the cell and its environment. Most small molecules cross the membrane by the general mechanisms shown schematically in Figure 2–5 and explained as follows:
-
Diffusion transports small, nonpolar molecules directly through the lipid bilayer. Lipophilic (fat-soluble) molecules diffuse through membranes readily, water very slowly.
-
Channels are multipass proteins forming transmembrane pores through which ions or small molecules pass selectively. Cells open and close specific channels for Na+, K+, Ca2+ and other ions in response to various physiological stimuli. Water molecules usually cross the plasma membrane through channel proteins called aquaporins.
-
Carriers are transmembrane proteins that bind small molecules and translocate them across the membrane via conformational changes.
Major mechanisms by which molecules cross membranes.

Lipophilic and some small, uncharged molecules can cross membranes by simple diffusion (a).
Most ions cross membranes in multipass proteins called channels (b) whose structures include transmembrane ion-specific pores.
Many other larger, water-soluble molecules require binding to sites on selective carrier proteins (c), which then change their conformations and release the molecule to the other side of the membrane.
Diffusion, channels and most carrier proteins translocate substances across membranes using only kinetic energy. In contrast, pumps are carrier proteins for active transport of ions or other solutes and require energy derived from ATP.
Diffusion, channels, and carrier proteins operate passively, allowing movement of substances across membranes down a concentration gradient due to its kinetic energy. In contrast, membrane pumps are enzymes engaged in active transport, utilizing energy from the hydrolysis of adenosine triphosphate (ATP) to move ions and other solutes across membranes, against often steep concentration gradients. Because they consume ATP pumps they are often referred to as ATPases.
These transport mechanisms are summarized with additional detail in Table 2–2.
Mechanisms of transport across the plasma membrane.
| Process | Type of Movement | Example |
|---|---|---|
| PASSIVE PROCESSES | Movement of substances down a concentration gradient due to the kinetic energy of the substance; no expenditure of cellular energy is required; continues until equilibrium is reached (if unopposed) | |
| Simple diffusion | Unassisted net movement of small, nonpolar substances down their concentration gradient across a selectively permeable membrane | Exchange of oxygen and carbon dioxide between blood and body tissues |
| Facilitated diffusion | Movement of ions and small, polar molecules down their concentration gradient; assisted across a selectively permeable membrane by a transport protein | |
| Channel-mediated | Movement of ion down its concentration gradient through a protein channel | Na+ moves through Na+ channel into cell |
| Carrier-mediated | Movement of small, polar molecule down its concentration gradient by a carrier protein | Transport of glucose into cells by glucose carrier |
| Osmosis | Diffusion of water across a selectively permeable membrane; direction is determined by relative solute concentrations; continues until equilibrium is reached | Solutes in blood in systemic capillaries “pulls” fluid from interstitial space back into the blood |
| ACTIVE PROCESSES | Movement of substances requires expenditure of cellular energy | |
| Active transport | Transport of ions or small molecules across the membrane against a concentration gradient by transmembrane protein pumps | |
| Primary | Movement of substance up its concentration gradient; powered directly by ATP | Ca2+ pumps transport Ca2+ out of the cell Na+/K+ pump moves Na+ out of cell and K+ into cell |
| Secondary | Movement of a substance up its concentration gradient is powered by harnessing the movement of a second substance (eg, Na+) down its concentration gradient | |
| Symport | Movement of substance up its concentration gradient in the same direction as Na+ | Na+/glucose transport |
| Antiport | Movement of substance up its concentration gradient in the opposite direction from Na+ | Na+/H+ transport |
| Vesicular transport | Vesicle formed or lost as material is brought into a cell or released from a cell | |
| Exocytosis | Bulk movement of substance out of the cell by fusion of secretory vesicles with the plasma membrane | Release of neurotransmitter by nerve cells |
| Endocytosis | Bulk movement of substances into the cell by vesicles forming at the plasma membrane | |
| Phagocytosis | Type of endocytosis in which vesicles are formed as particulate materials external to the cell are engulfed by pseudopodia | White blood cell engulfing a bacterium |
| Pinocytosis | Type of endocytosis in which vesicles are formed as interstital fluid is taken up by the cell | Formation of small vesicles in capillary wall to move substances |
| Receptor-mediated endocytosis | Type of endocytosis in which plasma membrane receptors first bind specific substances; receptor and bound substance then taken up by the cell | |
The cell membrane separates the interior of cells from the outside environment. The cell membrane is selectively permeable to ions and organic molecules and controls the movement of substances in and out of cells.
It consists of the phospholipid bilayer with embedded proteins.
Cell membranes are involved in a variety of cellular processes such as cell adhesion, ion conductivity and cell signalling and serve as the attachment surface for several extracellular structures, including the cell wall, glycocalyx, and intracellular cytoskeleton. Cell membranes can be artificially reassembled.
The limiting membranes that envelop all eukaryotic cells are made of phospholipids, cholesterol, proteins, and oligosaccharide chains covalently linked to phospholipid and protein molecules.
The cell membrane functions as a selective barrier regulating the passage of materials into and out of the cell and facilitating the transport of specific molecules. One important role of the cell membrane is to keep constant the ion content of cytoplasm, which differs from that of the extracellular fluid. Membranes also carry out a number of specific recognition and signaling functions, playing a key role in the interactions of the cell with its environment.The plasma membrane defines the outer limit of the cell. However a continuum exists between the interior of the cell and extracellular macromolecules. The plasma membrane contains proteins called integrins linked to both cytoplasmic protein filaments and ECM components. These linkages produce a continuous exchange of influences, in both directions, between the ECM and the cytoplasm.1
Membranes consists of a lipid bilayer and associated proteins. The plasma membrane has selective permeabilities and acts as a barrier, thereby maintaining differences in composition between the inside and outside of the cell. The selective permeabilities for substrates and ions are provided mainly by specific proteins named transporters and ion channels. The plasma membrane also exchanges material with the extracellular environment by exocytosis and endocytosis, and there are special areas of membrane structure—gap junctions—through which adjacent cells exchange material. In addition, the plasma membrane plays key roles in cell–cell interactions and in transmembrane signaling.
Membranes also form specialized compartments within the cell. Such intracellular membranes help shape many of the morphologically distinguishable structures (organelles), eg, mitochondria, ER, Golgi, secretory granules, lysosomes, and the nucleus. Membranes localize enzymes, function as integral elements in excitation-response coupling, and provide sites of energy transduction, such as in photosynthesis and oxidative phosphorylation.
Changes in membrane components can affect water balance and ion flux, and therefore many processes within the cell. Specific deficiencies or alterations of certain membrane components (eg, caused by mutations genes encoding membrane proteins) lead to a variety of diseases. In short, normal cellular function depends on normal membranes.
Membranes range from 7.5 to 10 nm in thickness and consequently are visible only in the electron microscope.
Membrane phospholipids are amphipathic, consisting of two nonpolar (hydrophobic or water-repelling) long-chain fatty acids linked to a charged polar (hydrophilic or water-attracting) head that bears a phosphate group.
Figure: Lipids in membrane structure.

(a) Membranes of animal cells have as their major lipid components phospholipids and cholesterol. A phospholipid is amphipathic, with a phosphate group charge on the polar head and two long, nonpolar fatty acid chains, which can be straight (saturated) or kinked (at an unsaturated bond). Membrane cholesterol is present in about the same amount as phospholipid.
(b) The amphipathic nature of phospholipids produces the bilayer structure of membranes as the charged (hydrophilic) polar heads spontaneously form each membrane surface, in direct contact with water, and the hydrophobic nonpolar fatty acid chains are buried in the membrane's middle, away from water. Cholesterol molecules are also amphipathic and are interspersed less evenly throughout the lipid bilayer; cholesterol affects the packing of the fatty acid chains, with a major effect on membrane fluidity. The outer layer of the cell membrane also contains glycolipids with extended carbohydrate chains.
Sectioned, osmium-fixed cell membrane may have a faint trilaminar appearance with the transmission electron microscope (TEM), showing two dark (electron-dense) lines enclosing a clear (electron-lucent) band. Reduced osmium is deposited on the hydrophilic phosphate groups present on each side of the internal region of fatty acid chains where osmium is not deposited. The "fuzzy" material on the outer surface of the membrane represents the glycocalyx of oligosaccharides of glycolipids and glycoproteins. X100,000.
Phospholipids are most stable when organized into a double layer (bilayer) with the hydrophobic fatty acid chains directed toward the middle away from water and the hydrophilic polar head groups facing the water. Molecules of cholesterol, a sterol lipid, insert at varying densities among the closely-packed phospholipid fatty acids, restricting their movement, and modulating the fluidity and movement of all membrane components. The phospholipids in each half of the bilayer are different. For example, in the well-studied membranes of red blood cells phosphatidylcholine and sphingomyelin are more abundant in the outer half, while phosphatidylserine and phosphatidylethanolamine are more concentrated in the inner layer. Some of the outer lipids, known as glycolipids, include oligosaccharide chains that extend outward from the cell surface and contribute to a delicate cell surface coating called the glycocalyx. With the transmission electron microscope (TEM) the cell membrane—and all other organellar membranes—may exhibit a trilaminar appearance after fixation in osmium tetroxide; osmium binding the polar heads of the phospholipids, the outer sugar chains, and associated membrane proteins produces the two dark outer lines enclosing the light band of osmium-free fatty acids.
Figure 2–2 Proteins associated with the membrane lipid bilayer.

(a) The fluid mosaic model of membrane structure emphasizes that the phospholipid bilayer of a membrane also contains proteins inserted in it or associated with its surface (peripheral proteins) and that many of these proteins move within the fluid lipid phase. Integral proteins are firmly embedded in the lipid layers; those that completely span the bilayer are called transmembrane proteins. Hydrophobic amino acids of these proteins interact with the hydrophobic fatty acid portions of the membrane lipids. Both the proteins and lipids may have externally exposed oligosaccharide chains.
(b) When cells are frozen and fractured (cryofracture), the lipid bilayer of membranes is often cleaved along the hydrophobic center. Splitting occurs along the line of weakness formed by the fatty acid tails of phospholipids. Electron microscopy of such cryofracture preparation replicas provides a useful method for studying membrane structures. Most of the protruding membrane particles seen (1) are proteins or aggregates of proteins that remain attached to the half of the membrane adjacent to the cytoplasm (P or protoplasmic face). Fewer particles are found attached to the outer half of the membrane (E or extracellular face). Each protein bulging on one surface has a corresponding depression (2) on the opposite surface.
- Although the plasma membrane defines the outer limit of the cell, a continuum exists between the interior of the cell and extracellular macromolecules. The plasma membrane contains proteins called integrins linked to both cytoplasmic protein filaments and ECM components. These linkages produce a continuous exchange of influences, in both directions, between the ECM and the cytoplasm.
- Membranes are dynamic structures. Lipids and certain proteins show rapid lateral diffusion. Flip–flop is very slow for lipids and nonexistent for proteins.
- The fluid mosaic model forms a useful basis for thinking about membrane structure.
- Membrane proteins are classified as integral if they are firmly embedded in the bilayer and as peripheral if they are attached to the outer or inner surface.
- The 20 or so membranes in a mammalian cell have different compositions and functions and they define compartments, or specialized environments, within the cell that have specific functions (eg, lysosomes).
- Certain hydrophobic molecules freely diffuse across membranes, but the movement of others is restricted because of their size or charge.
- Various passive and active (usually ATP-dependent) mechanisms are employed to maintain gradients of such molecules across different membranes.
- Certain solutes, eg, glucose, enter cells by facilitated diffusion along a downhill gradient from high to low concentration using specific carrier proteins (transporters).
- The major ATP-driven pumps are classified as P (phosphorylated), F (energy factors), V (vacuolar), and ABC transporters. Member of these classes include the Na+–K+-ATPase and the Ca2+ ATPase of the sarcoplasmic reticulum; the mt ATP synthase; the ATPase acidifying lysosomes; and the CFTR protein and the MDR-1 protein.
- Ligand- or voltage-gated ion channels are often employed to move charged molecules (Na+, K+, Ca2+, etc.) across membranes down their electrochemical gradients
- Large molecules can enter or leave cells through mechanisms such as endocytosis or exocytosis. These processes often require binding of the molecule to a receptor, which affords specificity to the process.
- Receptors may be integral components of membranes (particularly the plasma membrane). The interaction of a ligand with its receptor may not involve the movement of either into the cell, but the interaction results in the generation of a signal that influences intracellular processes (transmembrane signaling).
- Mutations that affect the structure of membrane proteins (receptors, transporters, ion channels, enzymes, and structural proteins) may cause diseases; examples include cystic fibrosis and familial hypercholesterolemia.
- Membranes are structures composed of lipids, proteins, and carbohydrate-containing molecules.
- The plasma membrane consists of a bilayer of phospholipids. This bilayer is formed by two sheets of phospholipids in which the hydrophilic polar head groups are directed away from each other and are exposed to the aqueous environment on the outer and inner surfaces of the membrane. The hydrophobic nonpolar tails of these molecules are oriented toward each other, in the direction of the center of the membrane.
Membrane phospholipids are amphipathic, consisting of two nonpolar (hydrophobic or water-repelling) long-chain fatty acids linked to a charged polar (hydrophilic or water-attracting) head that bears a phosphate group. Their hydrocarbon chains are oriented inward to the center of the bilayer to form a continuous hydrophobic phase and their hydrophilic heads oriented outward.
 (a) Membranes of animal cells have as their major lipid components phospholipids and cholesterol. A phospholipid is amphipathic, with a phosphate group charge on the polar head and two long, nonpolar fatty acid chains, which can be straight (saturated) or kinked (at an unsaturated bond). Membrane cholesterol is present in about the same amount as phospholipid.
(a) Membranes of animal cells have as their major lipid components phospholipids and cholesterol. A phospholipid is amphipathic, with a phosphate group charge on the polar head and two long, nonpolar fatty acid chains, which can be straight (saturated) or kinked (at an unsaturated bond). Membrane cholesterol is present in about the same amount as phospholipid.
(b) The amphipathic nature of phospholipids produces the bilayer structure of membranes as the charged (hydrophilic) polar heads spontaneously form each membrane surface, in direct contact with water, and the hydrophobic nonpolar fatty acid chains are buried in the membrane's middle, away from water. Cholesterol molecules are also amphipathic and are interspersed less evenly throughout the lipid bilayer; cholesterol affects the packing of the fatty acid chains, with a major effect on membrane fluidity. The outer layer of the cell membrane also contains glycolipids with extended carbohydrate chains.
Sectioned, osmium-fixed cell membrane may have a faint trilaminar appearance with the transmission electron microscope (TEM), showing two dark (electron-dense) lines enclosing a clear (electron-lucent) band. Reduced osmium is deposited on the hydrophilic phosphate groups present on each side of the internal region of fatty acid chains where osmium is not deposited. The "fuzzy" material on the outer surface of the membrane represents the glycocalyx of oligosaccharides of glycolipids and glycoproteins. X100,000.
Proteins associated with the membrane lipid bilayer.
 (a) The fluid mosaic model of membrane structure emphasizes that the phospholipid bilayer of a membrane also contains proteins inserted in it or associated with its surface (peripheral proteins) and that many of these proteins move within the fluid lipid phase. Integral proteins are firmly embedded in the lipid layers; those that completely span the bilayer are called transmembrane proteins. Hydrophobic amino acids of these proteins interact with the hydrophobic fatty acid portions of the membrane lipids. Both the proteins and lipids may have externally exposed oligosaccharide chains.
(a) The fluid mosaic model of membrane structure emphasizes that the phospholipid bilayer of a membrane also contains proteins inserted in it or associated with its surface (peripheral proteins) and that many of these proteins move within the fluid lipid phase. Integral proteins are firmly embedded in the lipid layers; those that completely span the bilayer are called transmembrane proteins. Hydrophobic amino acids of these proteins interact with the hydrophobic fatty acid portions of the membrane lipids. Both the proteins and lipids may have externally exposed oligosaccharide chains.
(b) When cells are frozen and fractured (cryofracture), the lipid bilayer of membranes is often cleaved along the hydrophobic center. Splitting occurs along the line of weakness formed by the fatty acid tails of phospholipids. Electron microscopy of such cryofracture preparation replicas provides a useful method for studying membrane structures. Most of the protruding membrane particles seen (1) are proteins or aggregates of proteins that remain attached to the half of the membrane adjacent to the cytoplasm (P or protoplasmic face). Fewer particles are found attached to the outer half of the membrane (E or extracellular face). Each protein bulging on one surface has a corresponding depression (2) on the opposite surface.
Molecules of cholesterol, a sterol lipid, insert at varying densities among the closely-packed phospholipid fatty acids, restricting their movement, and modulating the fluidity and movement of all membrane components. The phospholipids in each half of the bilayer are different. For example, in the well-studied membranes of red blood cells phosphatidylcholine and sphingomyelin are more abundant in the outer half, while phosphatidylserine and phosphatidylethanolamine are more concentrated in the inner layer. Some of the outer lipids, known as glycolipids, include oligosaccharide chains that extend outward from the cell surface and contribute to a delicate cell surface coating called the glycocalyx. With the transmission electron microscope (TEM) the cell membrane—and all other organellar membranes—may exhibit a trilaminar appearance after fixation in osmium tetroxide; osmium binding the polar heads of the phospholipids, the outer sugar chains, and associated membrane proteins produces the two dark outer lines enclosing the light band of osmium-free fatty acids.
Proteins are major constituents of membranes (~50% by weight in the plasma membrane). Integral proteins are directly incorporated within the lipid bilayer itself, whereas peripheral proteins exhibit a looser association with one of the two membrane surfaces, particularly the inner (Figure 2–2). The loosely bound peripheral proteins can be easily extracted from cell membranes with salt solutions, whereas integral proteins can be extracted only by using detergents to disrupt lipids. The polypeptide chains of many integral proteins span the membrane several times, from one side to the other, and are accordingly called multipass transmembrane proteins. Integration of the proteins within the lipid bilayer is mainly the result of hydrophobic interactions between the lipids and nonpolar amino acids present on the outer region of the proteins.
Freeze-fracture electron microscope studies of membranes show that parts of many integral proteins protrude from both the outer or inner membrane surface (Figure 2–2b). Like those of glycolipids, the carbohydrate moieties of glycoproteins project from the external surface of the plasma membrane and contribute to the glycocalyx (Figure 2–3). They are important components of proteins acting as receptors, which participate in important interactions such as cell adhesion, cell recognition, and the response to protein hormones. As with lipids, the distribution of membrane polypeptides is different in the two surfaces of the cell membranes. Therefore, all membranes in the cell are asymmetric.
The plasma membrane contains proteins called integrins linked to both cytoplasmic protein filaments and ECM components. These linkages produce a continuous exchange of influences, in both directions, between the ECM and the cytoplasm.


Membrane composition
The major chemical components of all membranes are lipids and proteins. Carbohydrates are associated mainly with plasma membranes, where they combine chemically with lipids, forming glycolipids, and with proteins, forming glycoproteins.
The outer side of the membrane is covered by a glycocalyx.
Although the plasma membrane defines the external limit of the cell, a continuum exists between the interior of the cell and extracellular macromolecules. The plasma membrane contains proteins called integrinsthat are linked to both cytoplasmic cytoskeletal filaments and extracellular matrix components. Through these linkages there is a constant exchange of influences, in both directions, between the extracellular matrix and the cytoplasm.

Membranes range from 7.5 to 10 nm in thickness and consequently are visible only in the electron microscope.
References:
1. http://www.ncbi.nlm.nih.gov/books/NBK21054/
2. http://www.johnkyrk.com/er.swf
With the Electron Microscope
Research Instrument:
1. http://sine.ni.com/cs/app/doc/p/id/cs-502
2. http://www.mathworks.nl/matlabcentral/newsreader/view_thread/292249
Cell membranes can be artificially reassembled.[4][5][6]
Quiz
1. Mescher AL. Mescher A.L. Mescher, Anthony L.Chapter 2. The Cytoplasm. In: Mescher AL. Mescher A.L. Ed. Anthony L. Mescher.eds. Junqueira's Basic Histology: Text & Atlas, 13e. New York, NY: McGraw-Hill; 2013. http://accessmedicine.mhmedical.com/content.aspx?bookid=574&Sectionid=42524588. Accessed March 20, 2015.
Tansmembrane Channels
Transmembrane channels, also called membrane channels, are pores within the lipid bilayer of the cell membrane. The channels are formed by large protein complexes that run across the membrane connecting the cytoplasm, to the extracellular matrix. Transmembrane channels are also found in the membranes of organelles including the nucleus, mitochondria and lysosome.
Transporters are membrane proteins that are present in all organisms. These proteins control the influx of essential nutrients and ions and the efflux of cellular waste, environmental toxins, drugs, and other xenobiotics. Consistent with their critical roles in cellular homeostasis, ∼2000 genes in the human genome, ∼7% of the total number of genes, code for transporters or transporter-related proteins. The functions of membrane transporters may be facilitated (equilibrative, not requiring energy) or active (requiring energy). In considering the transport of drugs, pharmacologists generally focus on transporters from two major superfamilies, ABC (ATP binding cassette) and SLC (solute carrier) transporters.
Most ABC proteins are primary active transporters, which rely on ATP hydrolysis to actively pump their substrates across membranes. There are 49 known genes for ABC proteins that can be grouped into seven subclasses or families (ABCA to ABCG) (Borst and Elferink, 2002). Among the best recognized transporters in the ABC superfamily are P-glycoprotein (P-gp, encoded by ABCB1, also termed MDR1) and the cystic fibrosis transmembrane regulator (CFTR, encoded by ABCC7).
The SLC superfamily includes genes that encode facilitated transporters and ion-coupled secondary active transporters that reside in various cell membranes. Forty-eight SLC families with ∼315 transporters have been identified in the human genome (Hediger, 2004). Many serve as drug targets or in drug absorption and disposition. Widely recognized SLC transporters include the serotonin (5-HT) and dopamine transporters (SERT, encoded by SLC6A4; DAT, encoded by SLC6A3).
Analysis of physical chemical evidence suggests that the involvement of transporters in the passage of drugs across biological membranes may be more the rule than the exception (Dobson and Kell, 2008). Drug-transporting proteins operate in pharmacokinetic and pharmacodynamic pathways, including pathways involved in both therapeutic and adverse effects (Figure 5–1).
Transmembrane channels differ from transporters and pumps in several ways; channels are not as selective as transporters and pumps, differentiating solutes primarily by size and ionic charge. Channels perform passive transport of materials also known as facilitated diffusion. Transporters can carry out either passive or active transfer of materials while pumps require energy to act.[3]
There are several modes by which membrane channels operate. The most common is the gated channel which requires a trigger, such as a change in membrane potential in voltage-gated channels, to unlock or lock the pore opening. Voltage-gated channels are critical to the production of an action potential in neurons resulting in a nerve impulse. A ligand-gated channel requires a chemical, such as a neurotransmitter, to activate the channel. Stress-gated channels require a mechanical force applied to the channel for opening. Aquaporins are dedicated channels for the movement of water across the hydrophobic interior of the cell membrane.[4]
Ion channels are a type of transmembrane channel responsible for the passive transport of positively charged ions (sodium, potassium, calcium, hydrogen and magnesium) and negatively charged ions (chloride) and, can be either gated or ligand-gated channels. One of the best studied ion channels is the potassium ion channel. The potassium ion channel can allow rapid movement of potassium ions while being selective against sodium. Using X-ray diffraction data and atomic model computations a likely structure of the channel consists of a number of protein alpha-helixes forming an hourglass shaped pore with the narrowest point halfway through the membrane's lipid bilayer. To move through the channel the potassium ions must shed their aqueous matrix and enter a selectivity filter composed of carbonyl oxygens. The potassium ions pass through one atom at a time along five different cation (positively charged ion) binding sites.[5]
Diseases caused by ion channel malfunctions include cystic fibrosis where the channel for the chloride ion will not open or is missing in the cells of the lungs, intestine, pancreas, liver and skin. The cells can no longer regulate salt and water concentrations resulting in the symptoms typical of the disease. Additional disorders resulting from malfunctions in ion channels include forms of epilepsy, cardiac arrhythmia, certain types of periodic paralysis and ataxia.[6]
Membrane composition
The major chemical components of all membranes are lipids and proteins. Intracellular membranes have a higher percentage of proteins than plasma membranes because enzymes which contain proteins exert their activity within organelles. Carbohydrates are associated mainly with plasma membranes, where they combine chemically with lipids, forming glycolipids, and with proteins, forming glycoproteins.
Lipids
The basic
References:
1. http://www.ncbi.nlm.nih.gov/books/NBK21054/
2. http://www.johnkyrk.com/er.swf
With the Electron Microscope
Research Instrument:
1. http://sine.ni.com/cs/app/doc/p/id/cs-502
2. http://www.mathworks.nl/matlabcentral/newsreader/view_thread/292249
Quiz
1. http://www.quia.com/quiz/1263142.html?AP_rand=1394007946
Cell wall membranes can include or exclude various molecules, and by controlling the movement of substances from one compartment to another, membranes exert a powerful influence on metabolic pathways. The plasma membrane also has an important role in cell-to-cell recognition. Other functions of the plasma membrane include cellular mobility and the maintenane of cellular shape.
References:
1. http://www.ncbi.nlm.nih.gov/books/NBK21054/
2. http://www.johnkyrk.com/er.swf
With the Electron Microscope
Research Instrument:
1. http://sine.ni.com/cs/app/doc/p/id/cs-502
2. http://www.mathworks.nl/matlabcentral/newsreader/view_thread/292249
Quiz
1. http://www.quia.com/quiz/1263142.html?AP_rand=1394007946
Cell membranes envelope eukaryotic cells. The plasma membrane (cell membrane) functions as a selectively permeable barrier regulating the passage of materials into and out of the cell and facilitating the transport of specific molecules. One important role of the cell membrane is to keep constant the ion content of cytoplasm, which differs from that of the extracellular fluid.
Membranes also carry out a number of specific recognition and signaling functions, playing a key role in the interactions of the cell with its environment.
Cell membranes are involved in a variety of cellular processes such as cell adhesion, ion conductivity and cell signaling, cell-to-cell recognition, cellular mobility and the maintenane of cellular shape and serve as the attachment surface for several extracellular structures, including the cell wall, glycocalyx, and intracellular cytoskeleton.
Individual lipid molecules in the bilayer vary according to the particular membrane and can move laterally and organize themselves with cholesterol (e.g., sphingolipids), endowing the membrane with fluidity, flexibility, organization, high electrical resistance, and relative impermeability to highly polar molecules. Membrane proteins embedded in the bilayer serve as structural anchors, receptors, ion channels, or transporters to transduce electrical or chemical signaling pathways and provide selective targets for drug actions.
Membranes are highly ordered and compartmented (Pinaud et al., 2009; Singer, 2004). These proteins may be associated with caveolin and sequestered within caveolae; they may be excluded from caveolae; or they may be organized in signaling domains rich in cholesterol and sphingolipid not containing caveolin or other scaffolding proteins (i.e., lipid rafts).
Questions?

{kind=link}
{kind=link}